| MadSci Network: Neuroscience |
The details of all the various neurotransmitter systems, e.g. anatomy,
receptors, responses, and interactions, are admittedly very complex.
But, after all, the number of classical neurotransmitters is limited. One
can safely assume that the more important ones have already been described.
Most of the classical NTs are either derived from dietary sources or
converted to the active compound by a few enzymatic reactions.
(These enzyme cascades have probably been well conserved because they were
subject to strong evolutionary pressures guaranteeing that these enzymes
function properly, especially when they perform tasks in other biochemical
pathways as well. After all, enzymes that don't do the job are usually
detrimental.)
Adding to the neurotransmitters, a plethora of neuroactive peptides, which numbers in the dozens, can act as so called neuromodulators or neurohormones (bringing about long term or long range changes in synaptic transmission or hormone secretion) or exert their effects directly, i.e. as a NT in certain neural pathways, e.g. substance P is involved in pain signalling. There is a great deal of diversity among these neuropeptides although they can be grouped into some families based on similarities in structure. Function can be conserved but there are cases where the function has changed during the evolution of organisms.
Recently even gaseous molecules, most notably nitric oxide (NO), have
been proven to act as neurotransmitters/-modulators.
I shouldn't forget to mention that organisms also have synapses where
electrical signals spread out between adjacent cells through specialized
structures, so called connexons, which are in effect ion channels.
Messenger molecules such as acetylcholine, amino acid transmitters,
biogenic amines (dopamine, etc.) have been around for some time and have
been detected in unicellular organisms. These transmitters underwent a
functional change while being structurally conserved and were coopted as
neurotransmitters in multicellular organisms. This also implies that
signalling within and between cells existed long before specialized nerve
cells evolved.
The diversity among receptors is indeed one reason for the wide range
of responses to a single neurotransmitter. There are (as of 1996) 5 GABA, 9
glutamate, 13 serotonin, and 11 different adrenergic receptors.
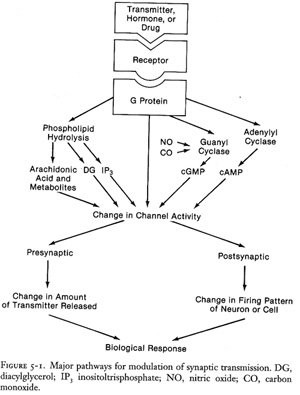
The more interesting part is though how receptors elicit cellular
responses. They can do this by either coupling directly to ion channels
(sodium, potassium, calcium, chloride) and thereby changing ion fluxes into
and out of the cell (excitation/inhibition), or they jump start various
second messenger systems mostly via G-proteins which again activate or
inhibit other proteins (e.g. ion channels, enzymes such as protein kinases,
etc.).
Either way the activity status of the cell changes. The information
about what a cell or cell population experiences is coded for in the
temporal and spatial (where and when) pattern of signals that impinge on
it.
In some cases the interaction between different receptors for the same neurotransmitter (even on the same cell) is crucial, e.g. in long term potentiation (cf. the glossary), where one type of glutamate receptor has to be activateds before another glutamate receptor type can act as a calcium channel.
The balance between neurotransmitters matters. Take for example
Parkinson's Disease and Chorea Huntington, disorders which
both affect the motor system. In normal individuals there seems to be a
delicate balance between the dopamine, acetylcholine, and GABA systems.
This balance is disrupted in:
a) Parkinson's Disease because dopamine levels in the substantia nigra
(literally, black substance, because dopamine is stored there in the form
of melanin, the pigment that gives you a tan) decrease severely,
b) Chorea H. because here certain cholinergic (acetylcholine containing)
and GABAergic neurons degenerate and die which results in low levels of
these NTs in brain areas involved in motor control.
Glossary:
G protein: class of proteins that undergo a change in activity after
exchanging cGDP (cyclic guanosine diphosphate) for cGTP (cyclic guanosine
triphosphate). The protein is usually active when GTP is bound.
LTP: fairly well explained at this Web
address.
References:
Try the links in the MadSci Library for more information on Neuroscience.