| MadSci Network: Genetics |
Thank you for the interesting question Catharine. You should know that your question sparked a considerable amount of debate among my colleagues, which is part of the reason why it has taken me as long as it has to answer. Most of our debate revolved around what exactly you meant by an "8x risk of genetic problems," because that figure seemed rather high. Based on studies of inbreeding the Japanese population, it seems like the actual risk of genetic defects due to these sort of matings is about 2x that of matings between unrelated individuals. In addition, I found an interesting paper looking at the effects of consanguinity on child mortality in Pakistan. That study found that the risk of infant mortality in the first 5 years was 1.2x greater for consanguinous families than for the rest of the population. References for these studies can be found at the bottom of this page.
Anyway, let's get on to addressing your question.
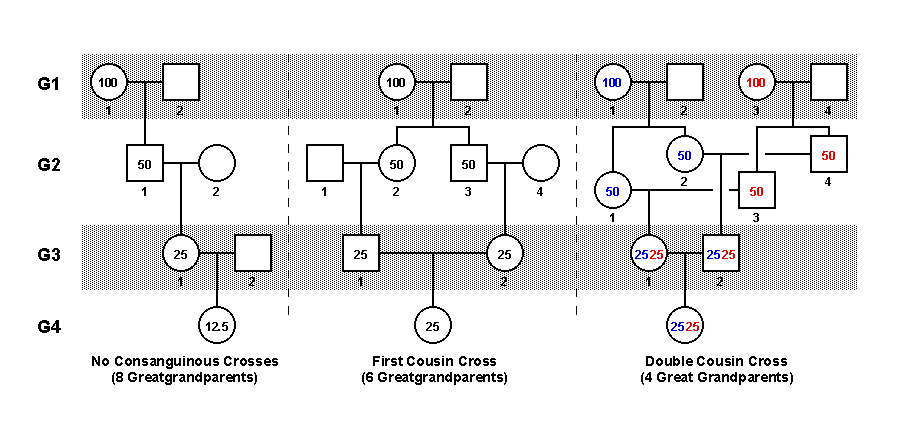
First off, lets take a look at what you mean by the offspring of double cousins. I've created the figure below, which describes three types of family pedigrees.
Figure #1
Let me explain this figure. The leftmost column of this figure indicates the four different generations in the pedigrees, G1- G4. Individuals in each pedigree are indicated with squares and circles -- circles for the women and squares for the men, and each individual in each generation of each pedigree has a number which identifies them as unique. For each of these pedigrees, we are going to be examining the genetic risks of the person in G4, the great-grandchild of the people in G1.
The leftmost section of the figure (labelled "No Consanguinous Crosses") is basically a subsection of a much larger pedigree in which the various people who are mating are unrelated to one another. Numbers 1 and 2 in G1 are both unrelated to one another, and both of the number 2s in G2 and G3 are unrelated as well.
The middle section depicts a mating of first cousins in G3, but as was the case with the left most section, 1 and 2 in G1 and 1 and 4 in G2 are unrelated.
The rightmost section depicts a mating of so-called double cousins in G3. These people are called double cousins because they are the offspring of a pair of brothers who married (I decided to keep the whole thing as wholesome as possible by making sure that everyone married everyone else :) a pair of sisters. However, the same conclusions would be true for a brother-sister pair if both of them married the siblings in another sister-brother pair. The nature of the siblings makes no difference.
Finally, I put numbers in some of the circles and squares. These represent the percent chance of having one copy of a particular allele of a particular gene. In the leftmost and middle sections, the numbers are black, but in the rightmost section, some numbers are red and some are blue. Each of these colors represents the chance of having a specific allele that is distinct from the allele represented by the other color.
What does it all mean?
In most cases, a person suffers from a genetic defect because they have inherited two copies of a defective allele for a particular gene (aka, they are homozygous for that allele). With two defective copies, the gene is not properly expressed, and an abnormality develops. People who have only one defective copy (allele) of a particular gene are said to be carriers for the particular defect that results when you inherit two copies. Now, it just so happens that each of us is a carrier of some sort of defective allele. We each have about 30,000 genes, and on average it is thought that each of us has about 5-10 defective alleles that we can pass on to the next generation. So, when we think about the increased risk of genetic defects due to inbreeding, we need to look at the chances of inheriting a particular defective allele. Please note however, that inheriting two copies of a so-called defective allele does not always yield a condition that is considered to be a "genetic defect". For example, colorblindness results from the inheritance of two defective genes for color vision, but is not really considered by the general public to be a genetic defect.
The pedigrees
The leftmost pedigree depicts a succession of unrelated matings. The way I have drawn it, it shows us the chance that the person in G4 will have inherited one copy of a defective allele that person number 1 in G1 had. Since we know that each of us has about 5-10 defective alleles, we can be sure that the great grandmother in G1 had at least one. So I filled in a 100 percent chance that she has one copy of that allele. For now, lets assume that no one else in the population has that allele, so we don't have to worry about all the number 2 people in the other generations.
As you can see, in each successive generation, the chance that someone will inherit the allele in question decreases by 50%, so that by the time we get to G4, the great granddaughter has a 12.5% (or 1 in 8) chance of having inherited one copy of the defective allele.
Now, lets look at the middle pedigree. Here, everything is the same as it was in the first pedigree until we have a mating in G3. There, because these individuals are first cousins, they each have a 25% chance of having one copy of the defective allele. So, the chance that the person in G4 will inherit one copy of this allele from them is one half of each of their chances of having the allele, added together. So you can see that her chance is the same as the chances of her parents. So, right off the bat we can see that the offspring of first cousin matings have 2x the chance of inheriting one copy of a particular allele.
In addition, because each of her parents has a 25% chance of having inherited that allele, the great granddaughter in G4 has a chance of inheriting 2 copies of the defective allele, with the resulting potential for genetic defects. The chance of this is half of the chance that her parents will have inherited the allele, squared (12.5% X 12.5%) or about 1.6%. This is much higher than the chance for the great granddaughter in the first pedigree, who had basically zero chance of inheriting two copies, because we stipulated that no one else in the population has that allele. We will revisit this again below with a more-realistic estimate. Bear with me.
Now lets look at the rightmost pedigree. Here, the person in G4 has only 4 great grandparents, where the other great granddaughters had more. Lets suppose that each of the great grandmothers has a different defective allele (red or blue), so that each of them has a 100% chance of having a defective allele, although they both don't have the same defective allele. When you follow the alleles down through the generations to G4, you see that the chance of having one copy of either the blue or red allele is the same in the great granddaughter in this pedigree as it was in the middle pedigree (25%), and it follows that the chance of getting two copies of a given allele is the same as well (1.6%). This is because, as far as an individual gene is concerned, there is no difference between a double cousin cross and a first cousin cross (as long as the great grandparents are unrelated). Follow the numbers yourself to make sure.
The interesting thing here is that while this is true on the level of an individual gene, the number of defective alleles that the great granddaughter in the rightmost pedigree has a 25% chance of inheriting is higher in the double cousin cross than it is in the first cousin cross. This is because the great granddaughter in the double cousin cross has only four great grandparents, while the great granddaughter in the first cousin cross has six great grandparents. So, the great granddaughter in the double cousin cross has a 25% chance of inheriting one copy of ANY of the defective alleles that her great grandarents had (and a 1.6% chance of inheriting 2 copies), while the great granddaughter in the first cousin cross only has a 25% chance of inheriting one copy (and a 1.6% chance of inheriing 2 copies) of the defective alleles that the two great grandparents at the top of the pedigree (2 / 6 of her total great grandparents) have. So, that basically doubles the number of defective alleles for which the offspring of double cousin crosses are at risk.
Things in the real world
Now, the preceding discussion supposed that each defective allele was unique to a particular great grandparent. However, in the real world, these alleles all exist in the population at a certain frequency, and we can use some of the tools of population genetics to predict the chances that a particular ancestor will have one copy of a particular allele (aka frequency of heterozygotes). With that information in mind, we can see how the probabilities that I discussed above are affected by allele frequency.
Thanks to the Hardy-Weinberg equation, we know that if a particular allele has a frequency that we will note as p in the population then the frequency of individuals who have only one copy of that allele will be 2 X p X (1 - p). So if a given defective allele is present in one of the great grandparents in any of the pedigrees, the chance that it will be present in any of the other members of the pedigree not descended from that grandparent is 2(p)(1-p). With that in mind, we can re-calculate the chance of inheriting two copies of a defective allele found in a grandparent for a variety of values of p, in both the unrelated and first / double cousin pedigrees. I have tabulated that data below.
| Homozygote frequency in G4 of |
 | Homozygote frequency in G4 of | ||
| Allele Frequency | Heterozygote Frequency | Non consanguinous pedigree | First / Double cousin pedigrees | |
| 0.5 | 0.5 | 0.078125000 | 0.09765625 | |
| 0.4 | 0.48 | 0.073200000 | 0.09302500 | |
| 0.3 | 0.42 | 0.059325000 | 0.07980625 | |
| 0.2 | 0.32 | 0.039200000 | 0.06002500 | |
| 0.1 | 0.18 | 0.017325000 | 0.03705625 | |
| 0.01 | 0.0198 | 0.001311008 | 0.017536381 | |
| 0.001 | 0.001998 | 0.000125624 | 0.015812874 | |
| 0.0001 | 0.00019998 | 0.000012506 | 0.015643754 | |
| 0.00001 | 0.000019998 | 0.000001250 | 0.015626875 |
Looking at this table, you can see several interesting trends. First off, you can see that the chance of being homozygous in G4 for a given great grandmother's deleterious allele in the first pediegree decreases in step with the decreasing frequency of the allele in the population. However, when you compare this to the chance of being homozygous in G4 for a given great grandparent's allele in either of the cousin-mating pedigrees, you can see that while this number varies to some extent for high frequency alleles, it never drops below the minimum value of about 1.6% that we calculated earlier. So, the offspring of any kind of first cousin mating have a base chance of homozygosity of about 1.6%, which can go up depending on the frequency of the deleterious allele in the general population.
If you assume that homozygosity for deleterious alleles is related to genetic defects, you can compare the frequency of homozygotes for the non-consanguinous-mating and cousin-mating pedigrees and try to make some sort of extrapolation of increased risk. For high frequency alleles, the risk for first / double cousin matings rises from 1.25x to 2x as the frequency of the defective allele drops, but once the allele becomes extremely rare (frequency of 1/ 10000 or less) the risk is increased many thousand fold.
Summary
Okay, so that was a lot. Let me summarize:
So, overall I would not expect to see a tremendous increase in the risk of genetic defects in the offspring of so-called double cousins when compared to the offspring of first cousins. The empirical evidence suggests that the increased risk for first cousin matings is about 2x that of the general population, and since the offspring of so-called double cousins have twice as many defective alleles to consider, I would predict that their risk would be elevated by a comparable degree.
Well, I know I was long winded, but I hope that this has helped answer your question, as well as helping you to understand some of the complex issues involved. Keep on asking questions!
Gulzar H. Shah, Michael B. Toney, and Brian L. Pitcher. (1997) CONSANGUINITY AND CHILD MORTALITY: THE RISK FACED BY FAMILIES IN PAKISTAN
Fujiki N, Kohli Y, Kato T, Hirayama M, Mutoh T, Nakanaga M, Tokuda A, Nakazaki S, Dochin M, Mano K. (1992) Genetic disease patterns in Japan: a review. Hum Biol 6:855-867.